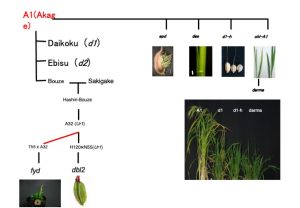
ニシヒカリ(Ur1): 北海道で定着したイネ在来種,赤毛から芒を失った坊主が琴似村で見出されました.芒は栽培に邪魔であること,坊主から早生系統が出現するなどしたことから,その後北海道の稲作の北進が加速していきました.いまからわずか130年程度前のことです.
さらに,坊主x魁から走坊主が育成されました.その栽培中に風連村にて走坊主中に発見され「風連坊主」(A32)と呼ばれました.その同型の穂型(枝梗湾曲,優性遺伝子Undulated-rachis1)を持つ永山村の「目黒栄糯」を母材としてN55が育成されています.高知大学の村井により,これら系統から佐藤繁俊先生(琉球大学)により育成されたT65sd1,Ur1を利用したニシヒカリ戻し交雑系統(戻し交雑を12回行っている)が育成されます.その後,自殖後代に9世代目において矮性個体が出現するなど高頻度で突然変異が観察されています.一部の系統では2-3mmの極矮小粒の表現型を示すキメラ性を検出しており,これらの後代から出現する変異体から原因遺伝子をクローニングする計画をすすめています.
A32系統自体からも変異体が出現するため,この系統と中国の系統,ACC848(岡彦一氏が維持していたOKAテスター系統)やタイの在来種Th5との交雑で高い転移活性が維持されています.これらの系統から出現した変異体もキメラ性を有することからトランスポゾンの活性が維持されています.この変異体遺伝子の単離も検討しています.

トランスポゾンが原因遺伝子から再転移した際にコード領域が復帰すると正常表現型に復帰します.このようなキメラ性も転移因子が関与していることを示唆しています.
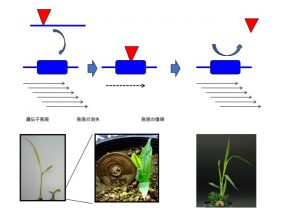
次世代シークエンサーを利用したゲノム情報から挿入を有するリードを単離して,その隣接領域からゲノム位置を推定できます.その情報をイネゲノム情報に照らし合わせて挿入サイトをゲノム上に位置づけられます.遺伝子情報もあるため,遺伝子内や制御領域と考えられる部分に挿入したものを候補として,プライマーを設定して単離します.変異体の分離と共分離(連鎖)するときに原因遺伝子であることが強く示唆されます.いま卒業研究で学生がこの手法で遺伝子単離を狙っています.
