トランスポゾンはマクリントックが提唱していらい,Class 1, Class 2にわけられます.Class 1は挿入しているマスター因子からの転写産物であるRNAを介して転移してDNAとして新たなサイトに挿入します.Class 2はDNAが挿入サイトから移動(転移)します.
Class 1 Retrotransposable elements related to retrovirus in structure
レトロウイルスと関連深いトランスポゾン
a RT (逆転写),GAG (RNAとりまくタンパク),Env (膜タンパク)
b Transcribed RNA transposes (SINE,LINE)
Class 2 DNA fragments transpose
Autonomous elements (自律因子ー転移するタンパクをコード)
Non-autonomous elements(非自律因子ー転移するには自律因子が必要)
当研究室では北海道の在来種で活性化した易変異性(えきへんいせい)について研究しています.赤毛はかって北海道の稲作の最初に定着した品種です.その自殖後代から数々の変異体が得られています.
背景:北海道における稲作は感温性品種である赤毛の選抜と加温による苗の育苗によることが知られている.在来種である赤毛の後代では大黒を始め,夷糯,坊主,坊主5号,坊主6号など形態や出穂性に変異を示す自然突然変異体が出現した.坊主と魁を交雑した走坊主から風連坊主が選抜され,優性突然変異であるUr1が多収に関わる優良変異であろうと研究が進められた.このUr1はAPO1と同座の変異であり,遺伝子上流の転写調節領域におけるnDart1の挿入が報告されている.一方,北海道大学の遺伝資源からA1(赤毛)の分譲を受け,弘前大学で自殖後代を展開し複数の自然変異体を得た. 本実験では,赤毛からの自殖後代に生じた変異の解析をすすめています.
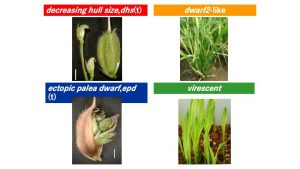
興味深い変異は下にしましたDECREASING HULL SIZE変異体です.劣性遺伝子により制御され,徐々に小穂(イネの花)のサイズが先端に向かって現象します.しかし,冬場に維持していてもそのような現象はみられません.この遺伝子の解析をすすめるとともに他に出現した変異体のクローニングを通して,赤毛がなぜ自殖後代で変異を生じさせることができるかについての遺伝的メカニズムについて調べています.

風連坊主(A32系統)からも変異体が出現しています.virecent変異は芒を失って葉緑素が減少します.この変異は劣性ですが発現が不安定です.この系統では既に優性変異,Ur1にトランスポゾンが挿入されていることが知られています(赤毛ではこのトランスポゾンの活性は認められていません).このトランスポゾン, nDartはAPO1に挿入した転移因子であり,Ur1はこのAPO1の制御因子であると考えられています.このようにUr1を持つことで易変性が維持されることから高知大学の村井先生が作成された系統,ニシヒカリ(Ur1)を利用した変異体の解析をすすめています.